
Nicotinamide Adenine Dinucleotide (NAD+) Peptide
Nicotinamide Adenine Dinucleotide (NAD+) is een geoxideerde vorm van NADH (Nicotinamide Adenine Dinucleotide Hydroxide). NAD+ is een component van de Elektronentransportketen (ETC), waarvan onderzoekers hebben gesuggereerd dat deze functioneert bij het transport van elektronen en daarmee energie binnen cellen. Er is ook verondersteld dat het peptide mogelijk fungeert als een mediator voor verschillende fysiologische processen, zoals post-translationele modificatie van eiwitten en activatie/deactivatie van sommige enzymen. Men gelooft dat het een cruciaal onderdeel is voor het behoud van communicatie tussen cellen.
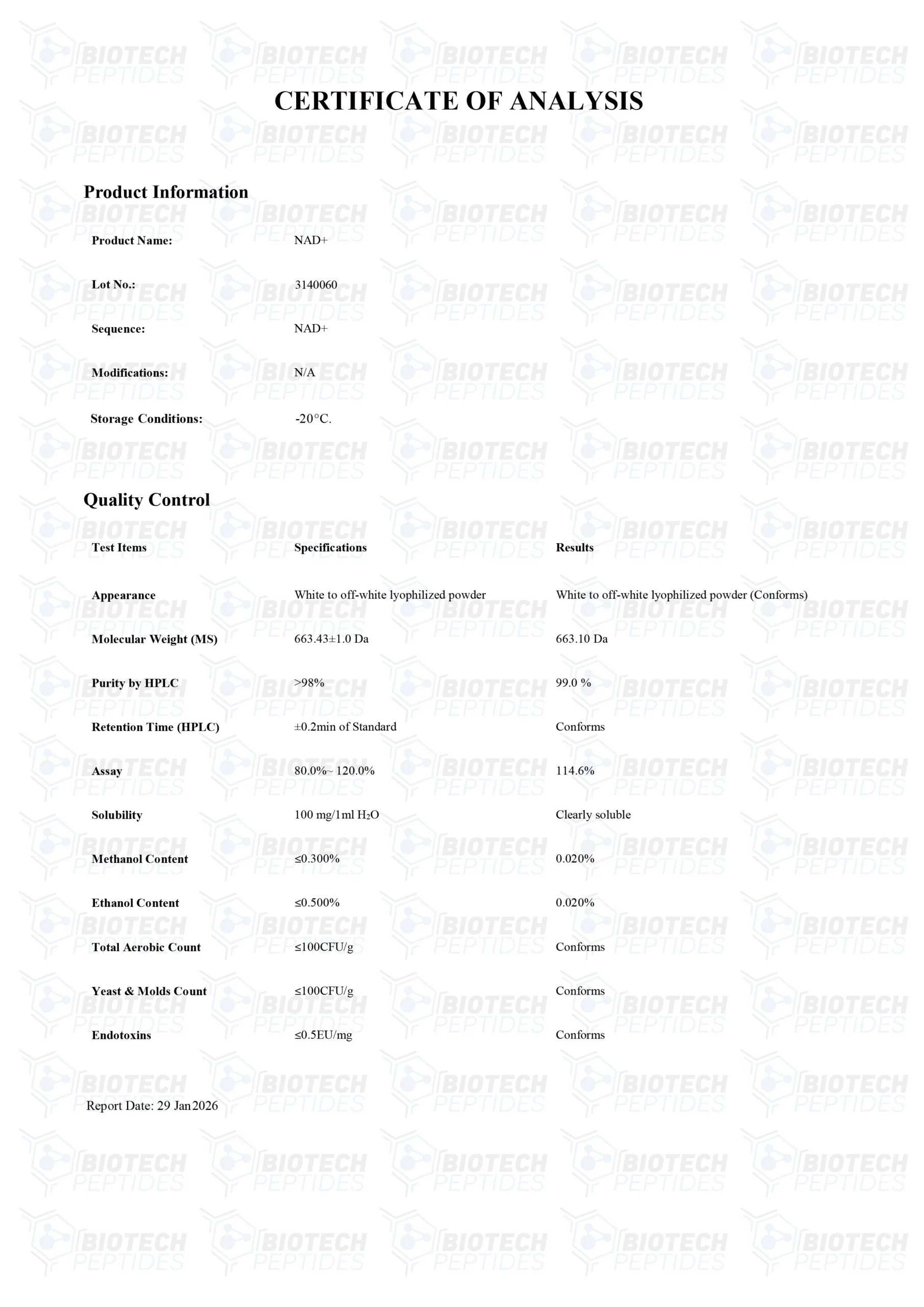
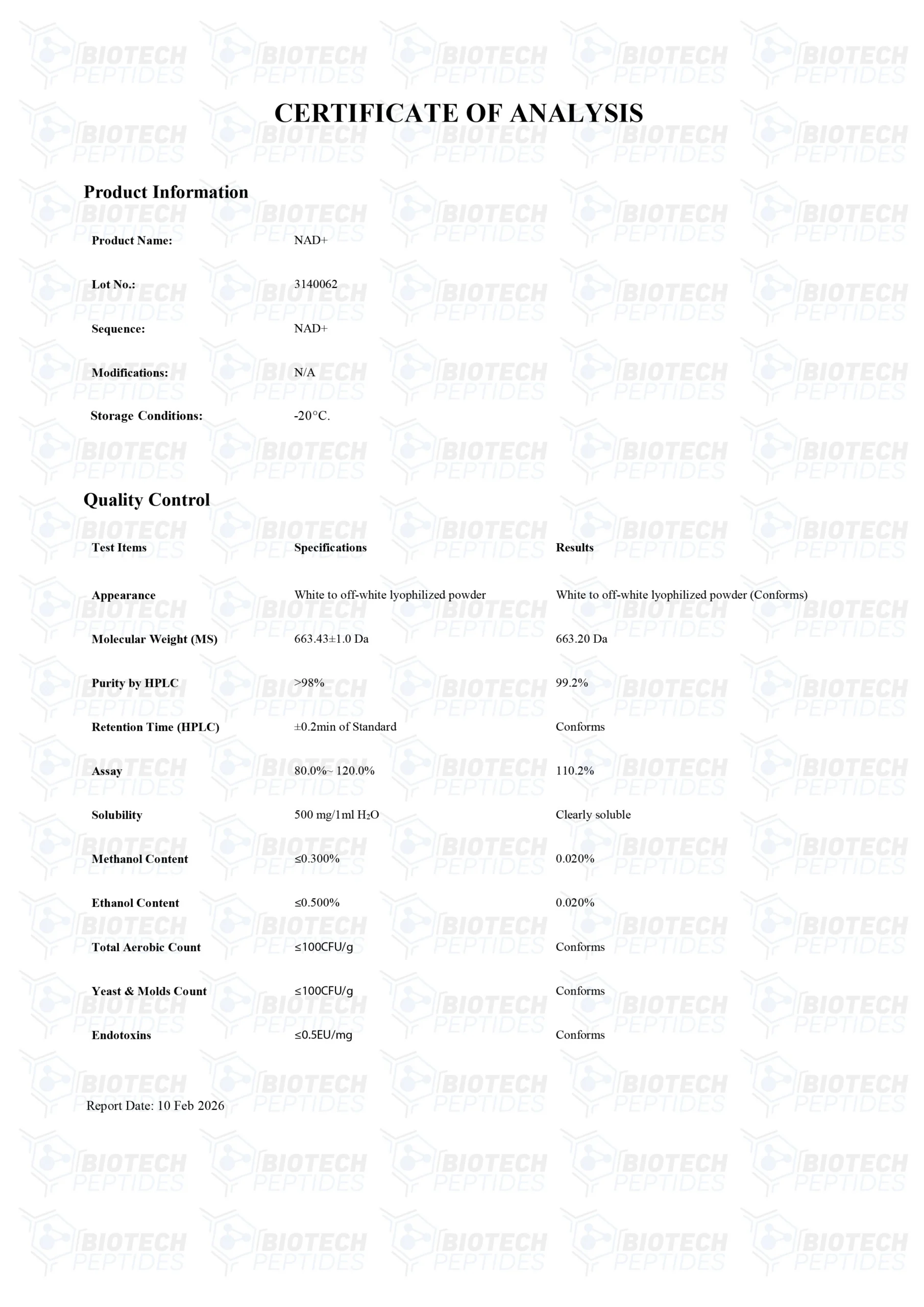
Specifications
MOLECULAIR GEWICHT: 663.43 g/mol
MOLECULAIRE FORMULE: C21H27N7O14P2
SYNONIEMEN: Nicotinamide Adenine Dinucleotide, Beta-NAD, NAD, Endopride
NAD+ Research
NAD+ and Cellular Aging
Mitochondriën dienen als een platform voor primaire metabolische functies zoals intracellulaire signalering en regulatie van aangeboren immuniteit. Deze processen lijken direct beïnvloed te worden door mitochondriale veroudering en veranderen uiteindelijk het cellulaire metabolisme, ontsteking en zelfs stamcelactiviteit.[1] Dit alles kan het tempo van weefselherstel na schade verminderen. Dit illustreert in welke mate mitochondriën betrokken zijn bij leeftijdsgerelateerde achteruitgang van cellulaire, weefsel- en orgaanfunctie. Onderzoekers zijn van mening dat zij door manipulatie van mitochondriale activiteit mogelijk het proces van cellulaire veroudering kunnen vertragen, stoppen of zelfs omkeren.
Een tekort aan NAD+ in de cel lijkt een pseudo-hypoxische toestand te veroorzaken, die de signalering binnen de celkern verstoort.[2] De wetenschappers suggereren dat “het verhogen van NAD+-niveaus in oude muizen de mitochondriale functie herstelt naar die van een jonge muis op een SIRT1-afhankelijke manier.” Het mechanisme achter deze eigenschap lijkt de activering van de SIRT1-functie te omvatten, waarbij een gen codeert voor een enzym genaamd Sirtuin-1 (NAD+-afhankelijke Deacetylase Sirtuin-1). Sirtuin-1 kan vervolgens mediatoren reguleren die betrokken zijn bij metabolisme, ontsteking en de levensduur van cellen.[3]
Sirtuïne-1 is een lid van een klasse eiwitten genaamd sirtuïnen, waarvan wordt verondersteld dat zij NAD+ als cofactor nodig hebben om mogelijke enzymatische activiteiten uit te voeren. Ze zijn mogelijk betrokken bij verschillende cellulaire processen, waaronder DNA-herstel, genexpressie en metabolische regulatie. Ze zijn in verband gebracht met cellulaire levensduur en levensspanne.[4]
NAD+ and Muscle Cell Function
De endogene afname van spiercelfunctie wordt geassocieerd met mitochondriale veroudering. Wetenschappers menen dat deze achteruitgang in twee stappen plaatsvindt. De eerste, tot op zekere hoogte omkeerbare stap, omvat verminderde expressie van mitochondriale genen. Deze genen lijken verantwoordelijk te zijn voor oxidatieve fosforylering (het proces waarmee mitochondriën energie produceren). De tweede, onomkeerbare stap bestaat uit een afname van genen die verantwoordelijk zijn voor oxidatieve fosforylering in de celkern.
Experimenten met muismodellen hebben melding gemaakt van een schijnbare omkering van stap 1 na blootstelling aan extra NAD+ voordat de cel naar stap 2 overgaat.[5] Het mechanisme achter deze interventie in mitochondriale veroudering kan het stabiliseren van de activiteit van Peroxisome Proliferator-activated Receptor Gamma Co-activator 1-alpha (PGC-1-alpha) omvatten. Studies hebben gesuggereerd dat de hierboven genoemde werking in de mitochondriën vergelijkbaar kan zijn met lichaamsbeweging op de mitochondriën van skeletspieren.[6]
NAD+ and Neurodegeneration
NAD+ is een cofactor die mogelijk een neuroprotectieve werking uitoefent.[7] Er is verondersteld dat dit kan gebeuren door ondersteuning van de mitochondriale functie en vermindering van de productie van reactieve oxidatieve stress (ROS). ROS is verantwoordelijk voor ontstekingsveranderingen die samenhangen met letsel en degeneratieve veranderingen die verband houden met cellulaire veroudering. Deze associatie vormt de basis voor bepaalde neurodegeneratieve ziekten, zoals Alzheimer, Huntington en Parkinson.
Onderzoek uitgevoerd bij muizen suggereerde het potentieel van NAD+ om te beschermen tegen progressieve motorische tekorten en het afsterven van dopamineproducerende cellen in de substantia nigra.[8] Volgens de onderzoekers: “Deze resultaten versterken de gunstige rol van NAD tegen parkinsonische neurodegeneratie in muismodellen van PD, leveren bewijs voor het potentieel van NAD voor de preventie van PD, en suggereren dat NAD pathologische veranderingen bij PD voorkomt door mitochondriale disfuncties te verminderen.” De onderzoeksresultaten impliceren dat hoewel NAD+ de symptomen niet lijkt te verlichten, het mogelijk de progressie van de ziekte van Parkinson kan vertragen of zelfs de ontwikkeling ervan kan beperken.
NAD+ and Inflammation
NAMPT is een enzym dat geassocieerd wordt met ontsteking. Het lijkt overmatig tot expressie te komen in bepaalde typen kankercellen. Een toename van de niveaus van NAMPT lijkt te correleren met NAD+-niveaus en vice versa.[9] De met NAMPT geassocieerde ontsteking lijkt voor te komen in kankercellen en onderzoeksmodellen van obesitas, type 2 diabetes en niet-alcoholische leververvetting. NAMPT kan een krachtige activator van ontsteking zijn, terwijl cellulaire ontstekingsniveaus mogelijk sterk afnemen na introductie van NAD+. De verandering van NAD+-niveaus zou ontstekingsroutes binnen cellen kunnen beïnvloeden, wat wijst op een mogelijke benadering om ontstekingsreacties op cellulair niveau te moduleren.
NAD+ and DNA Integrity
Onderzoek heeft onderzocht hoe NAD+ mogelijk de integriteit van DNA beschermt via de associatie met enzymen genaamd poly(ADP-ribose) polymerasen (PARPs).[10] Men denkt dat NAD+ dient als substraat voor PARP-enzymen, die mogelijk deelnemen aan DNA-herstelmechanismen door ADP-ribose (ADPr)-eenheden aan specifieke eiwitten te koppelen. Het enzym PARP-1, het eerste ontdekte lid van de PARP-familie, wordt verondersteld actief te worden tijdens DNA-schade door ketens van poly(ADP-ribose) (pADPr) aan eiwitten toe te voegen.
Deze pADPr-ketens kunnen fungeren als steigers die mogelijk DNA-hersteleiwitten aantrekken naar de locaties van schade en mogelijk helpen bij het losmaken van de chromatinestructuur om herstelprocessen te vergemakkelijken. De studie stelt ook voor dat pADPr aanvullende rollen binnen de celkern kan hebben onder normale omstandigheden, zoals het beïnvloeden van gentranscriptie, het veranderen van chromatine-architectuur en het onderhouden van telomeren — de beschermende uiteinden van chromosomen. Deze functies worden vermoedelijk gemedieerd door PARP-enzymen die NAD+ gebruiken om ADPr-modificaties op eiwitten te creëren. Verschillende PARP-enzymen kunnen samenwerken, waarbij sommige de toevoeging van enkele ADP-ribose-eenheden initiëren (een proces dat mono(ADP-ribosylering) wordt genoemd) en andere deze verlengen tot langere pADPr-ketens. Deze opeenvolgende modificatie kan de specificiteit en efficiëntie van DNA-herstelmechanismen ondersteunen.
Dit type onderzoek benadrukt dat andere enzymen die NAD+ verbruiken, zoals sirtuïnen, mogelijk ook bijdragen aan het behoud van DNA-integriteit. Sirtuïnen, geïllustreerd door het gisteiwit Sir2p, worden geassocieerd met gensilencing, chromosoomstabiliteit en cellulaire verouderingsprocessen. Specifiek wordt gerapporteerd dat de sirtuïne genaamd SIRT6 een enkele ADP-ribose-eenheid toevoegt aan PARP-1 als reactie op DNA-schade, wat vervolgens verdere toevoeging van poly(ADP-ribose)-ketens aan PARP-1 kan stimuleren. Dit suggereert dat er mogelijk een interactie bestaat tussen NAD+-afhankelijke signaleringsroutes bij het behoud van genomische stabiliteit.
NAD+ and Cell Survival
Als een cruciaal co-enzym dat een belangrijke rol speelt in cellulaire energiestofwisseling, kan NAD+ deelnemen aan verschillende signaalroutes die de overleving van cellen beïnvloeden. Onderzoekers suggereren dat onder omstandigheden van oxidatieve stress oxidatieve DNA-schade zich mogelijk ophoopt.[11] Deze ophoping kan het gevolg zijn van verhoogde aanvallen van reactieve zuurstofspecies (ROS) op DNA en een mogelijke afname van het DNA-herstelvermogen van de cel. De ophoping van DNA-laesies kan PARP-1 activeren, wat leidt tot uitputting van NAD+ en mogelijk resulteert in celdood.
In experimenten met neuronale celculturen die werden blootgesteld aan zuurstof-glucose-deprivatie (OGD) — een laboratoriummodel dat ischemisch letsel simuleert door de beschikbaarheid van zuurstof en glucose te verminderen — werd waargenomen dat het direct toevoegen van NAD+ vóór of na OGD de celdood leek te verminderen en DNA-schade leek te verlagen.[11] Deze beschermende werking leek afhankelijk te zijn van de concentratie NAD+ en het tijdstip van toediening. Er wordt verondersteld dat suppletie met NAD+ mogelijk nucleaire DNA-herstelactiviteiten herstelt door de fosforylering (toevoeging van fosfaatgroepen) van serine-aminozuren op essentiële enzymen die betrokken zijn bij base-excisieherstel (BER), zoals apurinisch/apyrimidinisch endonuclease 1 (APE1) en DNA-polymerase bèta (βpol), te verminderen. Het reactiveren van deze DNA-herstelmechanismen kan een belangrijke factor zijn die de neuroprotectieve werking van NAD+ medieert. Verdere observaties suggereren dat het aanvullen van NAD+ mogelijk de typische afname van BER-activiteiten vermindert die optreedt na OGD.
Het verlies van BER-functie in neuronen die aan OGD zijn blootgesteld, wordt vermoedelijk veroorzaakt door fosforylering op serine- en threonineresiduen van de snelheidsbepalende enzymen APE1 en βpol. NAD+ kan helpen de activiteiten van deze enzymen te herstellen door mogelijk proteïnekinasen (enzymen die fosfaatgroepen toevoegen) te remmen of fosfatasen (enzymen die fosfaatgroepen verwijderen) te activeren die hun fosforyleringstoestand reguleren. Experimenten waarbij de expressie van APE1 werd verminderd (knockdown), wezen erop dat dit de pro-overlevingsrelevantie van NAD+-suppletie aanzienlijk verminderde, wat impliceert dat de functionele integriteit van de BER-route cruciaal kan zijn voor de waargenomen neuroprotectie. Aangezien de knockdown van APE1 de pro-overlevingswerking echter niet volledig elimineerde, is het mogelijk dat NAD+ ook aanvullende overlevingsmechanismen activeert. Deze mechanismen kunnen onder meer het ondersteunen van de levering van substraten aan mitochondriën omvatten onder omstandigheden waarin NAD+ wordt verbruikt, waardoor energieproductie wordt bevorderd, of het activeren van NAD+-afhankelijke processen zoals SIRT-deacetylaseactiviteiten. Sirtuïnen kunnen de overleving van cellen beïnvloeden via chromatine-remodellering en onderdrukking van eiwitten die betrokken zijn bij apoptose (geprogrammeerde celdood).
Disclaimer
De genoemde producten zijn niet bedoeld voor menselijke of dierlijke consumptie. Research chemicaliën zijn uitsluitend bedoeld voor laboratoriumexperimenten en/of in-vitrotesten. Lichamelijke toediening van welke aard dan ook is strikt verboden volgens de wet. Alle aankopen zijn beperkt tot erkende onderzoekers en/of gekwalificeerde professionals. Alle informatie die in dit artikel wordt gedeeld, is uitsluitend bedoeld voor educatieve doeleinden.
References
Sun N, Youle RJ, Finkel T. The Mitochondrial Basis of Aging. Mol Cell. 2016 Mar 3;61(5):654-666. doi: 10.1016/j.molcel.2016.01.028. PMID: 26942670; PMCID: PMC4779179.
Gomes AP, Price NL, Ling AJ, Moslehi JJ, Montgomery MK, Rajman L, White JP, Teodoro JS, Wrann CD, Hubbard BP, Mercken EM, Palmeira CM, de Cabo R, Rolo AP, Turner N, Bell EL, Sinclair DA. Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 2013 Dec 19;155(7):1624-38. doi: 10.1016/j.cell.2013.11.037. PMID: 24360282; PMCID: PMC4076149.
Imai S, Guarente L. NAD+ and sirtuins in aging and disease. Trends Cell Biol. 2014 Aug;24(8):464-71. doi: 10.1016/j.tcb.2014.04.002. Epub 2014 Apr 29. PMID: 24786309; PMCID: PMC4112140.
Wątroba, M., Dudek, I., Skoda, M., Stangret, A., Rzodkiewicz, P., & Szukiewicz, D. (2017). Sirtuins, epigenetics and longevity. Ageing research reviews, 40, 11–19. https://doi.org/10.1016/j.arr.2017.08.001
Mendelsohn AR, Larrick JW. Partial reversal of skeletal muscle aging by restoration of normal NAD⁺ levels. Rejuvenation Res. 2014 Feb;17(1):62-9. doi: 10.1089/rej.2014.1546. PMID: 24410488.
Kang C, Chung E, Diffee G, Ji LL. Exercise training attenuates aging-associated mitochondrial dysfunction in rat skeletal muscle: role of PGC-1α. Exp Gerontol. 2013 Nov;48(11):1343-50. doi: 10.1016/j.exger.2013.08.004. Epub 2013 Aug 30. PMID: 23994518.
Matthews RT, Yang L, Browne S, Baik M, Beal MF. Coenzyme Q10 administration increases brain mitochondrial concentrations and exerts neuroprotective effects. Proc Natl Acad Sci U S A. 1998 Jul 21;95(15):8892-7. doi: 10.1073/pnas.95.15.8892. PMID: 9671775; PMCID: PMC21173.
Shan C, Gong YL, Zhuang QQ, Hou YF, Wang SM, Zhu Q, Huang GR, Tao B, Sun LH, Zhao HY, Li ST, Liu JM. Protective effects of β- nicotinamide adenine dinucleotide against motor deficits and dopaminergic neuronal damage in a mouse model of Parkinson’s disease. Prog Neuropsychopharmacol Biol Psychiatry. 2019 Aug 30;94:109670. doi: 10.1016/j.pnpbp.2019.109670. Epub 2019 Jun 17. PMID: 31220519.
Garten A, Schuster S, Penke M, Gorski T, de Giorgis T, Kiess W. Physiological and pathophysiological roles of NAMPT and NAD metabolism. Nat Rev Endocrinol. 2015 Sep;11(9):535-46. doi: 10.1038/nrendo.2015.117. Epub 2015 Jul 28. PMID: 26215259.
Leung A, Todorova T, Ando Y, Chang P. Poly(ADP-ribose) regulates post-transcriptional gene regulation in the cytoplasm. RNA Biol. 2012 May;9(5):542-8. doi: 10.4161/rna.19899. Epub 2012 May 1. PMID: 22531498; PMCID: PMC3495734.
Wang S, Xing Z, Vosler PS, Yin H, Li W, Zhang F, Signore AP, Stetler RA, Gao Y, Chen J. Cellular NAD replenishment confers marked neuroprotection against ischemic cell death: role of enhanced DNA repair. Stroke. 2008 Sep;39(9):2587-95. doi: 10.1161/STROKEAHA.107.509158. Epub 2008 Jul 10. PMID: 18617666; PMCID: PMC2743302.
Alle bestellingen worden dezelfde dag verzonden indien geplaatst vóór 12:00 PST.
Dit product is uitsluitend bestemd voor onderzoek-/laboratoriumgebruik. Menselijk of dierlijk gebruik en/of consumptie is strikt verboden volgens de wet. Alleen gekwalificeerde en erkende professionals mogen deze producten hanteren. Alle informatie gevonden op Biotech Peptiden is uitsluitend bedoeld voor educatieve doeleinden. Raadpleeg onze algemene voorwaarden voor meer details.
Dr. Usman
Dr. Usman (BSc, MBBS, MaRCP) voltooide zijn studie geneeskunde aan het Royal College of Physicians in Londen. Hij is een fervent onderzoeker met meer dan 30 publicaties in internationaal erkende peer-reviewed tijdschriften. Dr. Usman heeft gewerkt als onderzoeker en medisch consultant voor gerenommeerde farmaceutische bedrijven zoals Johnson & Johnson en Sanofi.



Reviews
There are no reviews yet.