
GHRP-6 Peptide
GHRP-6 peptide is een hexapeptide bestaande uit zes aminozuren en wordt door onderzoekers geclassificeerd als een groeihormoon-secretagoog (GHS), waarvan wordt gespeculeerd dat deze de afgifte van groeihormoon (GH) uit de cellen van de hypofysevoorkwab kan bevorderen. Het kan deze werking mogelijk bereiken door op te treden als een agonist van de ghrelinereceptor en behoort tot de ghreline-analogen die in de afgelopen decennia zijn ontwikkeld. Onderzoekers hebben gesuggereerd dat de ghrelinereceptor aanwezig is in de cellen van de hypofysevoorkwab. Aangezien de activatie ervan lijkt te resulteren in de synthese van groeihormoon, worden deze receptoren ook wel “groeihormoon-secretagoogreceptoren” (GHS-Rs) genoemd. Omdat wordt verondersteld dat ghreline invloed heeft op de stimulatie van het hongerhormoon, wordt ook aangenomen dat GHRP-6 vergelijkbare effecten kan uitoefenen en de voedselinname kan stimuleren. Daarnaast is gesuggereerd dat het een positieve invloed heeft op hartspiercellen, zelfs in gevallen van fibrose, en mogelijk positieve neurologische effecten kan hebben in experimentele settings.
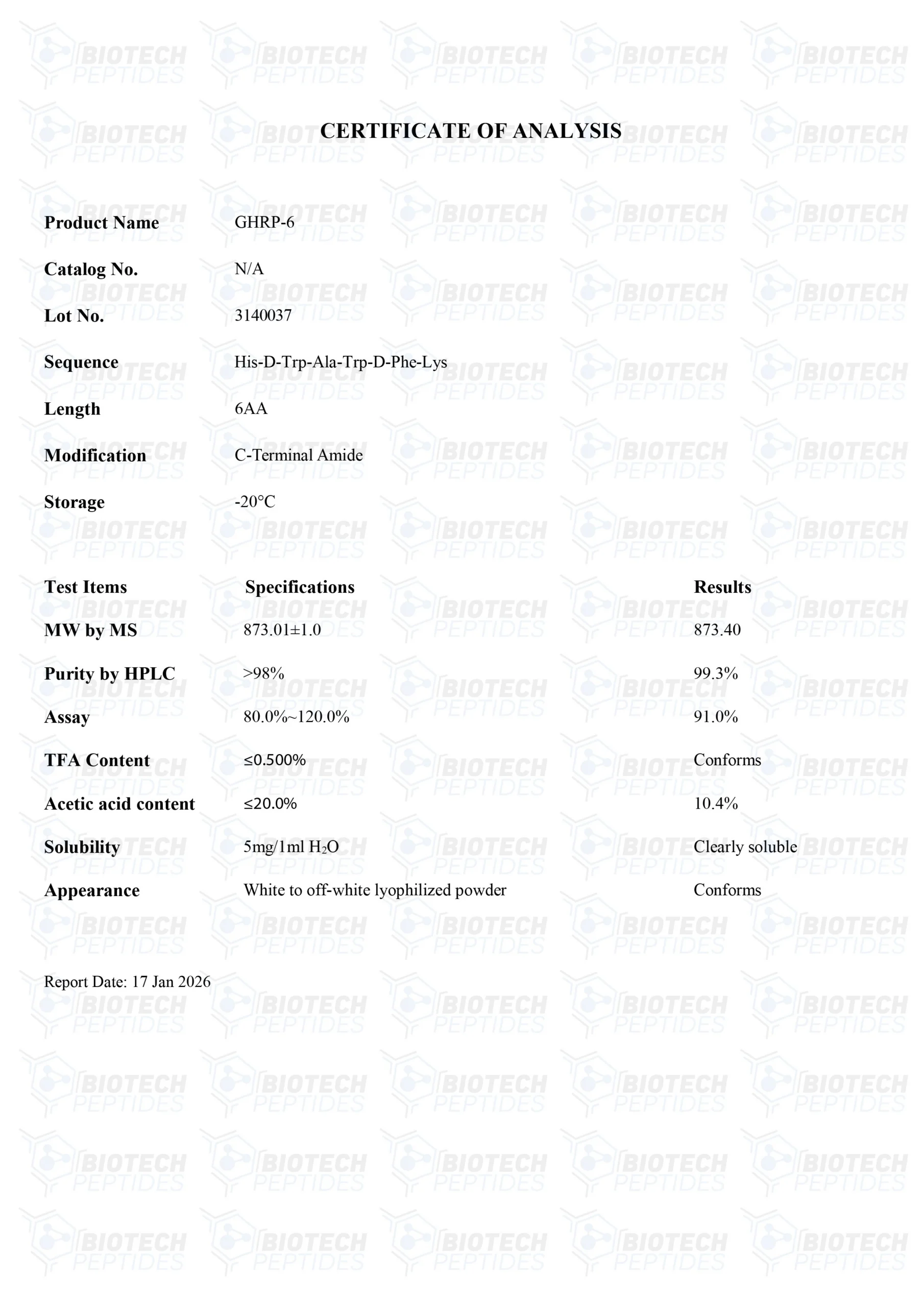
Specificaties
Moleculaire Formule: C46H56N12O6
Moleculair Gewicht: 873.032 g/mol
Sequentie: His-D-Trp-Ala-Trp-D-Phe-Lys
GHRP-6 Peptide Onderzoek
GHRP-6 en de GHS-receptoren
Na binding aan de GHS-receptoren activeert GHRP-6 waarschijnlijk een cascade van intracellulaire signaalprocessen die als cruciaal worden beschouwd voor de secretie van GH.[1] Een van de primaire routes die door GHRP-6 kan worden geactiveerd, betreft mogelijk de stimulatie van Gq-eiwitsubtypes. Deze stimulatie zou kunnen leiden tot activatie van fosfolipase C (PLC). Na activatie faciliteert PLC de hydrolyse van fosfatidylinositol 4,5-bisfosfaat (PIP2) in twee second messengers: inositoltrifosfaat (IP3) en diacylglycerol (DAG). De vrijgave van IP3 in het cytoplasma kan essentieel zijn voor het verhogen van intracellulaire calciumspiegels. IP3 lijkt dit te bereiken door binding aan receptoren op het endoplasmatisch reticulum (ER), waardoor calciumionen (Ca2+) uit deze intracellulaire opslagplaatsen vrijkomen. De stijging van cytosolisch calcium wordt beschouwd als een cruciaal signaal dat de fusie van GH-bevattende blaasjes met het plasmamembraan bevordert, wat leidt tot de afgifte van groeihormoon via exocytose. Tegelijkertijd lijkt DAG proteïnekinase C (PKC) te activeren, een belangrijk signaalmolecuul in talrijke cellulaire processen, waaronder hormoonsecretie. De activatie van PKC ondersteunt verder het proces van blaasjesfusie en GH-afgifte.
Onderzoekers hebben een mogelijke stijging van groeihormoonconcentraties (GH) waargenomen na blootstelling aan GHRP-6. De gegevens geven aan dat het gemiddelde piekniveau van GH ongeveer 15,7 nanogram per milliliter (ng/ml) bedroeg. Verder werd de cumulatieve gemiddelde hoeveelheid GH die gedurende de eerste 90 minuten van het onderzoek werd uitgescheiden, geregistreerd op 674 ng/ml.[2] In een afzonderlijke studie werden de effecten van GHRP-6 vergeleken met een controleverbinding. Deze analyse suggereert voorlopig dat de invloed van GHRP-6 correleerde met een afgifte van 15,4 ng/ml GH, in tegenstelling tot de controlegroep, die een niveau van 5,5 ng/ml vertoonde. Deze observaties impliceren een mogelijk verband tussen blootstelling aan GHRP-6 en verhoogde GH-spiegels.[3]
GHRP-6 en de CD36-receptoren
Er wordt verondersteld dat GHRP-6, een groeihormoonafgevend peptide, mogelijk kan interageren met andere receptorsites dan die welke specifiek gerelateerd zijn aan ghreline, bekend als GHS-Rs. Er bestaat een mogelijkheid dat deze aanvullende receptoren CD36-receptoren omvatten.[4] CD36-receptoren, betrokken bij een reeks biologische processen, zouden kunnen bijdragen aan het lipidenmetabolisme. Men denkt dat deze functie wordt gemedieerd via hun rol als scavengerreceptoren, die helpen bij de opname en het transport van lipiden door het organisme. Verder kunnen CD36-receptoren een rol spelen in het beïnvloeden van immuunreacties. Dit kan met name relevant zijn bij activiteiten zoals fagocytose, waarbij cellen schadelijke deeltjes opnemen, en ontsteking, een cruciale reactie van het immuunsysteem op infectie en letsel. Daarnaast zijn CD36-receptoren in verband gebracht met angiogenese, een complex proces waarbij nieuwe bloedvaten groeien uit reeds bestaande bloedvaten, wat essentieel wordt geacht voor weefselherstel en ontwikkeling.
GHRP-6 en Neurologische Disfunctie
Een studie uitgevoerd in 2018 benadrukte de prevalentie van ghrelinereceptoren in de substantia nigra, een deel van de hersenen dat wordt beschouwd als betrokken bij de pathologie van de ziekte van Parkinson.[1] Verder merkten de onderzoekers op dat zij “een dramatische afname vonden in de expressie van GHSR in PD-specifieke geïnduceerde pluripotente stamcel (iPSC)-afgeleide dopaminerge (DAnergische) neuronen gegenereerd uit [onderzoeksmodellen] met mutaties in het parkin-gen (PARK2).” Genetisch gepredisponeerde gevallen zouden een significante vermindering van ghrelinereceptoren in hun substantia nigra vertonen. Er werd waargenomen dat genetisch gemoduleerde ratten symptomen van Parkinson kunnen vertonen wanneer een antagonist aanwezig is. Wetenschappers veronderstellen dat het peptide dat geassocieerd is met receptoren in de substantia nigra mogelijk neuronale apoptose kan verminderen en zo het ontstaan van deze ziekte kan vertragen of verlichten.
GHRP-6 Peptide en Hersenceldood
De rol van het GHRP-6-peptide bij het verbeteren van beroertes is onderzocht in diermodellen. Blootstelling aan GHRP-6 kan mogelijk de verminderde bloedtoevoer naar hersenweefsel na een beroerte beperken en kan betrokken zijn bij geheugenherstel, wat tijdens een beroerte in gevaar kan komen.[2] Op moleculair niveau kunnen het peptide en zijn analogen apoptose van neuronen van het centrale zenuwstelsel voorkomen, waardoor genetische herprogrammering en ontsteking mogelijk worden verminderd.
GHRP-6 en Cognitieve Functie
Wetenschappers onderzoeken de rol van milde spierspanning in cognitieve functies die leren en geheugenvorming omvatten. Er is verondersteld dat spieractiviteit dergelijke functies kan ondersteunen. Het precieze mechanisme is echter onduidelijk gebleven. Aanvankelijk suggereerden onderzoekers dat fysieke activiteit een verbeterde bloedcirculatie in de hersenen faciliteerde en daardoor mogelijk de productie van groeihormoon verhoogde. Studies bij knaagdiermodellen hebben gesuggereerd dat GHRP-6 de overdracht van kortetermijngeheugen naar langetermijnopslag kan ondersteunen.[3] Belangrijke wetenschappelijke observaties suggereren verder een actieve rol van ghreline/GHRP-6 bij ruimtelijke leertaken.
GHRP-6 en Hartweefsel
Het GHRP-6-peptide zou mogelijk vrije-radicaal-gemedieerde cytotoxiciteit van hartcellen in varkensmodellen kunnen remmen.[4] De onderzoekers merkten op: “niveaus van oxidatieve stressmarkers suggereerden dat GHRP6 myocardiale schade voorkwam via een afname van reactieve zuurstofsoorten en door behoud van antioxidatieve afweersystemen.” Onderzoek op dit gebied is nog gaande.
GHRP-6 en Voortplanting
Studies bij mannelijke ratten hebben een positieve correlatie verondersteld van ghrelinereceptoren in het centrale zenuwstelsel bij het moduleren van opwinding en paringsimpulsen. GHRP-6-peptide en zijn gemodificeerde tegenhanger (waarvan wordt gesuggereerd dat deze de ghrelinereceptor antagoniseert) kunnen invloed hebben op hersengebieden die beloningszoekend gedrag faciliteren.[5] Er zijn ook gegevens die suggereren dat ghreline invloed kan hebben op stemming. Het peptide en zijn analogen lijken hersenfuncties te beïnvloeden die verband houden met stemming, secretie van stresshormonen en beloningsgedrag in muismodellen.[6]
GHRP-6 en Fibrose
Onderzoekers van GHRP-6 veronderstellen dat het peptide kan bijdragen aan celoverleving door geprogrammeerde celdood (apoptose) te verminderen. Het peptide is geassocieerd met de CD36-receptor, en onderzoekers suggereren dat het de groei van bloedvaten kan stimuleren, met name in beschadigd weefsel. Experimenten met GHRP-6-peptide en ratmodellen suggereren dat het natuurlijke herstelprocessen kan versnellen. Het lijkt de vorming van extracellulaire matrixeiwitten zoals collageen en de correcte organisatie van weefsel rond een litteken te ondersteunen, waardoor fibrose wordt verminderd. Hypertrofische littekens, zoals keloïden, kunnen ontstaan door onjuiste afzetting van matrixeiwitten, en dit proces kan mogelijk worden verbeterd door GHRP-6.[7]
Wetenschappers suggereren dat GHRP-6 dit afwijkende proces kan blokkeren.[8] In een recent experimenteel onderzoek met knaagdieren werd waargenomen dat blootstelling aan GHRP-6 gedurende een periode van 60 dagen mogelijk de ernst van leverfibrose verminderde.[13] Deze studie noteerde een daling in de niveaus van verschillende fibrogene markers, waaronder transformerende groeifactor-beta (TGF-β) en connective tissue growth factor (CTGF). Beide factoren lijken een cruciale rol te spelen in de ontwikkeling en progressie van fibreus weefsel. Specifiek gaf het onderzoek aan dat de fibrotische gebieden en nodulariteit in de lever respectievelijk met naar schatting 75% en meer dan 60% waren afgenomen. Dergelijke voorlopige gegevens suggereren dat GHRP-6 mogelijk kan helpen fibrose te beperken en herstelprocessen in experimentele settings kan ondersteunen.
GHRP-6 en Cellulaire Apoptose
Onderzoek heeft de mogelijke werking van GHRP-6 op neuronale cellen onderzocht, specifiek via zijn rol in het versterken van GH-productie en het vervolgens verhogen van niveaus van insulineachtige groeifactor-1 (IGF-1) in gerichte gebieden. IGF-1, structureel vergelijkbaar met insuline, fungeert als het belangrijkste anabole mechanisme van GH en faciliteert hypertrofie en mogelijk celdeling en ontwikkeling. Eerste observaties suggereren dat GHRP-6 de messenger RNA (mRNA)-expressie van IGF-1 kan verhogen binnen specifieke hersengebieden, waaronder de hypothalamus, het cerebellum en de hippocampus. Een dergelijke toename werd echter niet waargenomen in de cerebrale cortex. Deze selectieve verhoging wijst erop dat GHRP-6, via GH, de synthese van IGF-1 in specifieke neurale gebieden kan stimuleren. Bovendien evalueerde het onderzoek de expressie van de IGF-1-receptor en insulineachtige groeifactor-bindend eiwit 2 (IGFBP-2). IGFBP-2 reguleert de biologische beschikbaarheid van IGF-1 door eraan te binden, hoewel de studie geen significante veranderingen in hun expressie registreerde na blootstelling aan het peptide. Er werd echter fosforylering waargenomen van proteïnekinase B (Akt) en de Bcl-2-geassocieerde death promoter (BAD) in gebieden met verhoogd IGF-1 mRNA, wat mogelijk aangeeft dat GH en GHRP-6 cellulaire overlevingsroutes kunnen activeren als reactie op deze groeifactoren. BAD, een lid van de Bcl-2-eiwitfamilie, is cruciaal bij het reguleren van cellulaire overleving en dood. Daarentegen neemt Akt deel aan een breed spectrum van cellulaire processen, waaronder metabolisme, apoptose, groei, transcriptie en celmigratie. Daarnaast werd een toename van Bcl-2, een antiapoptotisch eiwit, waargenomen in dezelfde gebieden, wat duidt op een mogelijke neiging richting celoverleving in plaats van geprogrammeerde celdood. Daarentegen vertoonden de niveaus van het proapoptotische eiwit Bax geen verandering. Deze bevindingen suggereren voorlopig dat GHRP-6 bepaalde neuroprotectieve mechanismen in de hersenen kan beïnvloeden.
Disclaimer: De genoemde producten zijn niet bedoeld voor menselijke of dierlijke consumptie. Research chemicaliën zijn uitsluitend bedoeld voor laboratoriumexperimenten en/of in-vitrotesten. Elke vorm van lichamelijke toediening is wettelijk strikt verboden. Alle aankopen zijn beperkt tot erkende onderzoekers en/of gekwalificeerde professionals. Alle informatie in dit artikel is uitsluitend bedoeld voor educatieve doeleinden.
Referenties
Mousseaux D, Le Gallic L, Ryan J, Oiry C, Gagne D, Fehrentz JA, Galleyrand JC, Martinez J. Regulation of ERK1/2 activity by ghrelin-activated growth hormone secretagogue receptor 1A involves a PLC/PKCvarepsilon pathway. Br J Pharmacol. 2006 Jun;148(3):350-65. doi: 10.1038/sj.bjp.0706727. PMID: 16582936; PMCID: PMC1751558.
Cordido F, Peñalva A, Dieguez C, Casanueva FF. Massive growth hormone (GH) discharge in obese subjects after the combined administration of GH-releasing hormone and GHRP-6: evidence for a marked somatotroph secretory capability in obesity. J Clin Endocrinol Metab. 1993 Apr;76(4):819-23. doi: 10.1210/jcem.76.4.8473389. PMID: 8473389.
Frieboes RM, Murck H, Maier P, Schier T, Holsboer F, Steiger A. Growth hormone-releasing peptide-6 stimulates sleep, growth hormone, ACTH and cortisol release in normal man. Neuroendocrinology. 1995 May;61(5):584-9. doi: 10.1159/000126883. PMID: 7617137.
Demers, A., McNicoll, N., Febbraio, M., Servant, M., Marleau, S., Silverstein, R., & Ong, H. (2004). Identification of the growth hormone-releasing peptide binding site in CD36: a photoaffinity cross-linking study. The Biochemical journal, 382(Pt 2), 417–424. https://doi.org/10.1042/BJ20040036
Suda, Y., Kuzumaki, N., Sone, T., Narita, M., Tanaka, K., Hamada, Y., Iwasawa, C., Shibasaki, M., Maekawa, A., Matsuo, M., Akamatsu, W., Hattori, N., Okano, H., & Narita, M. (2018). Down-regulation of ghrelin receptors on dopaminergic neurons in the substantia nigra contributes to Parkinson’s disease-like motor dysfunction. Molecular brain, 11(1), 6. https://doi.org/10.1186/s13041-018-0349-8
Subirós, N., Pérez-Saad, H. M., Berlanga, J. A., Aldana, L., García-Illera, G., Gibson, C. L., & García-Del-Barco, D. (2016). Assessment of dose-effect and therapeutic time window in preclinical studies of rhEGF and GHRP-6 coadministration for stroke therapy. Neurological research, 38(3), 187–195. https://doi.org/10.1179/1743132815Y.0000000089
Huang, C. C., Chou, D., Yeh, C. M., & Hsu, K. S. (2016). Acute food deprivation enhances fear extinction but inhibits long-term depression in the lateral amygdala via ghrelin signaling. Neuropharmacology, 101, 36–45. https://doi.org/10.1016/j.neuropharm.2015.09.018
Berlanga, J., Cibrian, D., Guevara, L., Dominguez, H., Alba, J. S., Seralena, A., Guillén, G., López-Mola, E., López-Saura, P., Rodriguez, A., Perez, B., Garcia, D., & Vispo, N. S. (2007). Growth-hormone-releasing peptide 6 (GHRP6) prevents oxidant cytotoxicity and reduces myocardial necrosis in a model of acute myocardial infarction. Clinical science (London, England : 1979), 112(4), 241–250. https://doi.org/10.1042/CS20060103
Hyland, L., Rosenbaum, S., Edwards, A., Palacios, D., Graham, M. D., Pfaus, J. G., Woodside, B., & Abizaid, A. (2018). Central ghrelin receptor stimulation modulates sex motivation in male rats in a site dependent manner. Hormones and behavior, 97, 56–66. https://doi.org/10.1016/j.yhbeh.2017.10.012
Huang, H. J., Chen, X. R., Han, Q. Q., Wang, J., Pilot, A., Yu, R., Liu,Q., Li, B., Wu, G. C., Wang, Y. Q., & Yu, J. (2019). The protective effects of Ghrelin/GHSR on hippocampal neurogenesis in CUMS mice. Neuropharmacology, 155, 31–43. https://doi.org/10.1016/j.neuropharm.2019.05.013
Mendoza Marí, Y., Fernández Mayola, M., Aguilera Barreto, A., García Ojalvo, A., Bermúdez Alvarez, Y., Mir Benítez, A. J., & Berlanga Acosta, J. (2016). Growth Hormone-Releasing Peptide 6 Enhances the Healing Process and Improves the Esthetic Outcome of the Wounds. Plastic surgery international, 2016, 4361702. https://doi.org/10.1155/2016/4361702
Fernández-Mayola, M., Betancourt, L., Molina-Kautzman, A., Palomares, S., Mendoza-Marí, Y., Ugarte-Moreno, D., Aguilera-Barreto, A., Bermúdez-Álvarez, Y., Besada, V., González, L. J., García-Ojalvo, A., Mir-Benítez, A. J., Urquiza-Rodríguez, A., & Berlanga-Acosta, J. (2018). Growth hormone-releasing peptide 6 prevents cutaneous hypertrophic scarring: early mechanistic data from a proteome study. International wound journal, 15(4), 538–546. https://doi.org/10.1111/iwj.12895
Berlanga-Acosta, J., Vázquez-Blomquist, D., Cibrián, D., Mendoza, Y., Ochagavía, M. E., Miranda, J., … & Guillén-Nieto, G. E. (2012). Growth Hormone Releasing Peptide 6 (GHRP6) reduces liver fibrosis in CCl4 chronically intoxicated rats. Biotecnología Aplicada, 29(2), 60-72.
Frago LM, Pañeda C, Dickson SL, Hewson AK, Argente J, Chowen JA. Growth hormone (GH) and GH-releasing peptide-6 increase brain insulin-like growth factor-I expression and activate intracellular signaling pathways involved in neuroprotection. Endocrinology. 2002 Oct;143(10):4113-22. doi: 10.1210/en.2002-220261. PMID: 12239123.
Alle bestellingen worden dezelfde dag verzonden indien geplaatst vóór 12:00 PST.
Dit product is uitsluitend bedoeld voor onderzoek/laboratoriumgebruik. Menselijk of dierlijk gebruik en/of consumptie is wettelijk strikt verboden. Alleen gekwalificeerde en erkende professionals mogen deze producten hanteren. Alle informatie op Biotech Peptiden is uitsluitend bedoeld voor educatieve doeleinden. Raadpleeg onze algemene voorwaarden voor meer details.
Dr. Usman
Dr. Usman (BSc, MBBS, MaRCP) voltooide zijn studie geneeskunde aan het Royal College of Physicians in Londen. Hij is een gepassioneerd onderzoeker met meer dan 30 publicaties in internationaal erkende peer-reviewed tijdschriften. Dr. Usman heeft gewerkt als onderzoeker en medisch consultant voor gerenommeerde farmaceutische bedrijven zoals Johnson & Johnson en Sanofi.



Reviews
There are no reviews yet.