
PE-22-28 Peptide
PE-22-28 is een synthetische variant van het natuurlijk voorkomende peptide Spadin. Spadin is een afgescheiden peptide dat wordt verkregen uit Sortiline. Het lijkt te functioneren als een antagonist van de TREK-1 (TWIK-gerelateerd kaliumkanaal) receptor, een twee-porie kaliumkanaal dat is geïdentificeerd als een potentieel doelwit binnen depressieonderzoek en als mogelijke regulator van neurogenese. Studies bij muizen suggereren dat verwijdering van de TREK-1 receptor hen resistent kan maken tegen depressief gedrag en bijbehorende chemische onevenwichtigheden in de hersenen.[1] PE-22-28 vertegenwoordigt een kortere synthetische Spadin-analoog van zeven aminozuren met mogelijk hogere werkzaamheid en stabiliteit. Onderzoek in diermodellen is nog gaande.
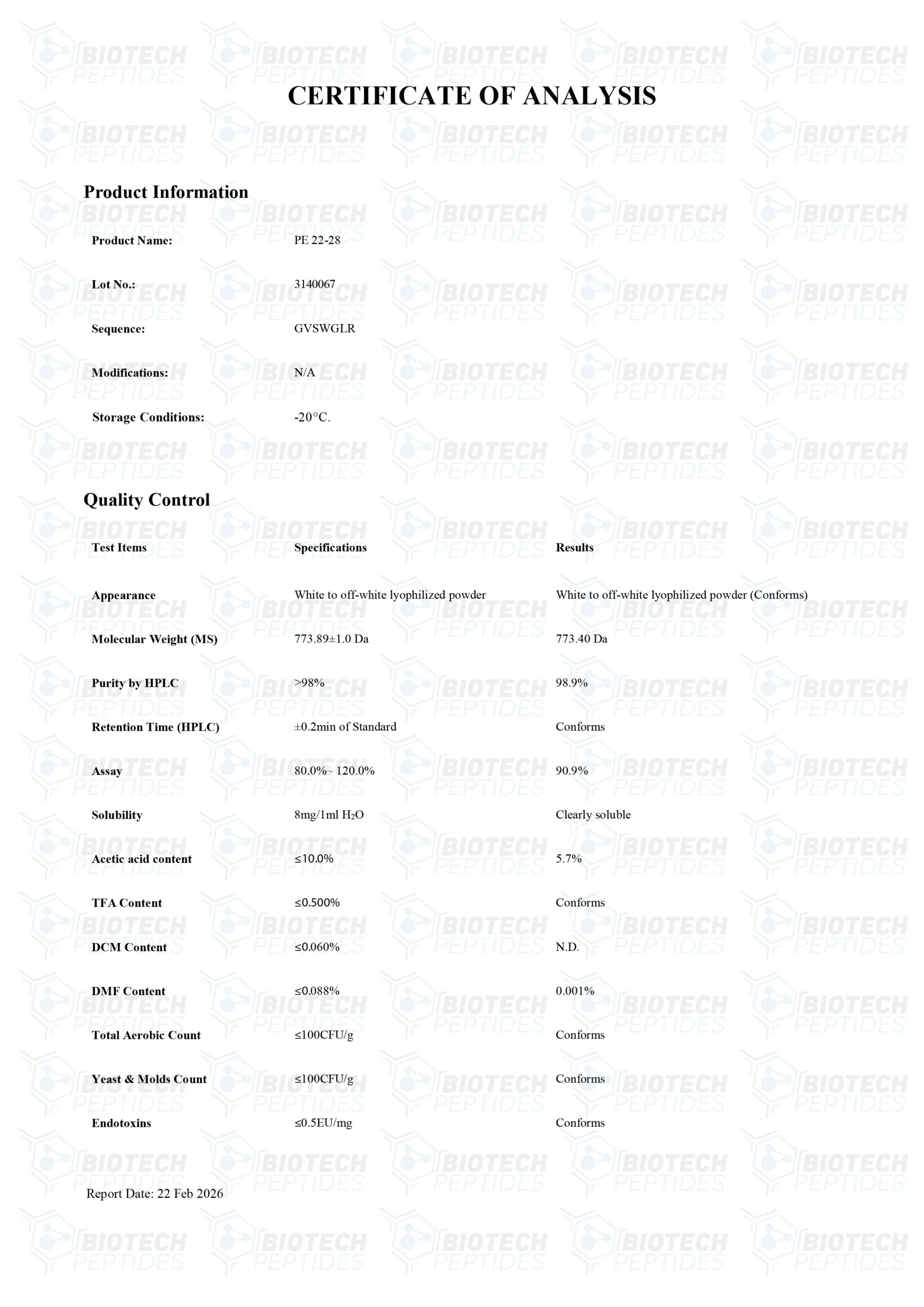
Specifications
Andere bekende namen: Spadin Analog, PE2228, TREK-1 Antagonist, PE 22 28, Sortilin Derivative, PE 2228
Sequentie: GVSWGLR
Moleculaire Formule: C35H55N11O9
Moleculair Gewicht: 773.8947 g/mol
PE-22-28 Research
Helaas is onderzoek naar PE-22-28 zeer beperkt vanwege een gebrek aan experimenten. Aangezien PE-22-28 een kortere versie van Spadin is met verondersteld verhoogde affiniteit en stabiliteit, hebben we hieronder de eigenschappen van Spadin uitgelicht, waarvan verwacht wordt dat ze ook van toepassing kunnen zijn op PE-22-28.
PE-22-28 and TREK-1 Receptors
TREK-1 is een verwante receptor van Spadin en onderzoekers suggereren daarom dat dit mogelijk ook geldt voor PE-22-28.[1] Het is een twee-porie kaliumkanaal dat wordt gereguleerd door verschillende moleculen en wordt beschouwd als een regulator van neuronale prikkelbaarheid.[2] TREK-1 lijkt overvloedig aanwezig te zijn in de prefrontale cortex, de amygdala en de hippocampus. Door de prikkelbaarheid van neuronen te verminderen, kan TREK-1 mogelijk bescherming bieden tegen excitotoxiciteit. Onderzoekers suggereren dat Spadin potentieel heeft om neurogene depressie, gereguleerd door het TREK-1 kanaal, te verlichten. Spadin kan mogelijk het verlies van hippocampusvolume omkeren door neurogenese te stimuleren.[3,4] Gezien de overeenkomsten tussen de potentiële werking van Spadin en PE-22-28, kan worden verondersteld dat een vergelijkbare hypothese ook voor PE-22-28 geldt.
PE-22-28 and BDNF
Er wordt verondersteld dat PE-22-28 mogelijk interacteert met BDNF, gebaseerd op onderzoek dat een dergelijke interactie tussen Spadin en BDNF suggereert. Brain-Derived Neurotrophic Factor (BDNF) is een eiwit dat mogelijk een rol speelt in de overleving, groei en instandhouding van neuronen en sterk betrokken is bij synaptische plasticiteit. Een studie suggereert dat Spadin mogelijk een rol speelt bij het moduleren van BDNF-niveaus, wat mogelijk bijdraagt aan de veronderstelde antidepressieve werking.[5] Spadin kan mogelijk indirect invloed uitoefenen op BDNF-routes, mogelijk via de impact op serotonine (5-HT) neurotransmissie en de regulatie van TREK-1 kanaalactiviteit. NTSR3/Sortiline, waaruit Spadin wordt afgeleid, wordt ook verondersteld interactie te hebben met neurotrofinen zoals pro-BDNF, die cruciaal worden geacht voor neurotrofische regulatie, celoverleving en geprogrammeerde celdood (apoptose). Hoewel beschikbaar onderzoek geen directe metingen van BDNF-niveaus of gerelateerde routes bevat, suggereert het dat verder onderzoek naar hoe Spadin BDNF op zowel mRNA- als eiwitniveau kan beïnvloeden, waardevolle inzichten kan bieden. Evenzo wordt verwacht dat PE-22-28 vergelijkbare of grotere potentie heeft op BDNF, maar onderzoek moet dit nog onderzoeken.
PE-22-28 Peptide and Neurogenesis
PE-22-28 lijkt neurogenese te bevorderen binnen een korte tijdsduur. Studies bij muizen suggereren dat PE-22-28 het potentieel heeft om neurogenese en synaptogenese al binnen vier dagen experimentatie te verhogen.[6] CREB (cAMP response element-binding protein) is een transcriptiefactor die wordt geassocieerd met neuronale plasticiteit, geheugenvorming en de ontwikkeling van ruimtelijk geheugen.[7] CREB lijkt niet alleen noodzakelijk te zijn voor de groei van neuronen, maar ook voor hun bescherming. De analyse suggereert dat kortdurende blootstelling aan Spadin leidt tot een duidelijke toename van gefosforyleerde CREB (pCREB)-niveaus, die ongeveer vier keer hoger werden waargenomen dan in placebo-blootgestelde controles. Deze toename in fosforylering wijst waarschijnlijk op CREB-activatie, ondersteund door Western blot-resultaten, die de actieve vorm van CREB toonden terwijl totale CREB-niveaus onveranderd bleven. Daarnaast werd een opvallende colocalisatie van pCREB met doublecortin (DCX) waargenomen, een eiwit dat vaak geassocieerd wordt met neuronale voorlopercellen. Deze colocalisatie impliceert dat CREB-activatie mogelijk nauw betrokken is bij het proces van neurogenese, met name met invloed op neuronale in plaats van gliale cellen. Daarom wordt verondersteld dat Spadin, mogelijk via snelle CREB-activatie, zowel de omvang als snelheid van hippocampale neurogenese aanzienlijk kan bevorderen.[5] Bijgevolg is het denkbaar dat PE-22-28 vergelijkbare effecten op neurogenese kan uitoefenen via vergelijkbare mechanismen.
Bevindingen suggereren ook dat Spadin mogelijk het aantal bromodeoxyuridine (BrdU)-positieve cellen in de hippocampus verhoogt in vergelijking met placebo, wat kan wijzen op een toename in de vorming van nieuwe neuronen, waarschijnlijk afkomstig van delende voorlopercellen. BrdU, een thymidine-analoog, wordt ingebouwd in het DNA van cellen tijdens de S-fase van de celcyclus en dient daardoor als indicator van celproliferatie. De schijnbaar snelle stijging van BrdU-positieve cellen binnen 4 dagen na blootstelling aan Spadin suggereert een versnelde activering van neurogene processen. Er wordt opnieuw verondersteld dat PE-22-28 op vergelijkbare wijze neurogenese via deze routes kan beïnvloeden.
Verder kunnen Spadin en analogen zoals PE-22-28 enkele van de nadelen missen die zijn waargenomen in modellen waarin TREK-1 kanalen volledig afwezig zijn. Onderzoek in diermodellen suggereert dat het verwijderen van het TREK-1 kanaal desastreus kan zijn voor het organisme. In eerdere muismodellen verhoogde uitschakeling van TREK-1 de kans op epileptische activiteit aanzienlijk en verminderde het het normale vermogen van dit twee-porie kaliumkanaal om neuronen tegen excitotoxiciteit te beschermen. Daarom was het verrassend dat noch Spadin noch PE-22-28 de epileptische activiteit na deze experimenten leek te verhogen. Nog interessanter is dat onderzoekers rapporteerden dat muizen blootgesteld aan Spadin resistenter leken tegen het ontwikkelen van gegeneraliseerde aanvallen. Onderzoek in diermodellen is nog gaande.
PE-22-28 Peptide and Serotonin
Zoals vermeld kan Spadin serotonine (5-HT) neurotransmissie moduleren via interactie met het TREK-1 kaliumkanaal, wat de vraag oproept of PE-22-28 vergelijkbaar potentieel kan hebben. Er wordt verondersteld dat de werking van Spadin de remming van TREK-1 omvat, wat leidt tot een verhoging van de vuursnelheid van 5-HT neuronen in de Dorsale Raphe Kern (DRN), een centraal knooppunt voor serotoninesignalering in de hersenen. Deze mogelijke blokkade kan de remmende feedback verminderen die normaal wordt gemedieerd door 5-HT1A autoreceptoren, wat op zijn beurt serotonine-neurotransmissie zou kunnen versterken.[5]
Verder suggereert aanvullend onderzoek dat de effecten van Spadin en serotonine receptoragonisten additief en onafhankelijk gemedieerd kunnen zijn.[8] Opmerkelijk is dat wanneer een antagonist van mGluR2/3 (metabotrope glutamaatreceptoren 2 en 3) wordt toegepast, dit de impact van Spadin lijkt te verminderen, wat impliceert dat de invloed van Spadin gekoppeld kan zijn aan TREK-1 kanalen binnen de mediale prefrontale cortex (mPFC) en hun interactie met mGluR2/3 receptoren.
Daarnaast geven immunohistochemische analyses aan dat Spadin, in combinatie met de selectieve serotonineagonist RS 67333, de expressie van Zif268 in de DRN kan veranderen, een marker gerelateerd aan neuronale activiteit. Zowel Spadin als RS 67333 afzonderlijk verhoogden het aantal Zif268-positieve neuronen, maar hun combinatie leidde tot een aanzienlijk grotere toename, wat wijst op een sterke depolariserende werking op een subgroep van DRN-neuronen. Interessant genoeg verhoogde de mGluR2/3 antagonist LY 341495 de vuursnelheid van DRN serotonine-neuronen. Dit effect werd echter niet waargenomen na een elektrolytische laesie in de mPFC, wat de mogelijke rol benadrukt van mPFC TREK-1 kanalen, mogelijk gekoppeld aan mGluR2/3 receptoren, bij het moduleren van deze activiteit. Aanvullende bevindingen uit fluorescentiemicroscopie, waarbij de Fura2-AM kleurstof werd gebruikt om intracellulaire calcium (Ca2+)-niveaus in gekweekte corticale neuronen te meten, toonden aan dat de combinatie van Spadin met LY 341495 of RS 67333 de intracellulaire Ca2+-niveaus sterker verhoogde dan blootstelling aan één van beide afzonderlijk. De meest uitgesproken stijging werd waargenomen bij de combinatie van Spadin en RS 67333. Dit effect lijkt afhankelijk te zijn van serotoninereceptoractivatie.
Er kan slechts worden verondersteld dat PE-22-28, als krachtige Spadin-analoog, vergelijkbare interacties kan vertonen met mGluR2/3 receptoren en serotonine-neuronen, mogelijk overeenkomend met de effecten die zijn waargenomen bij Spadin.
PE-22-28 Peptide and Muscle Function
Er bestaat enig onderzoek dat suggereert dat TREK-1 een rol speelt in het vermogen van spieren om te reageren op mechanische stimulatie.[9] Wetenschappers merken op dat “toepassing van negatieve druk op celgebonden patches (-20 mmHg) een 19-voudige toename veroorzaakte in de open waarschijnlijkheid (NPo) van … TREK-1 kanalen.” In het bijzonder lijkt blokkade van TREK-1 de contractiliteit in spierweefsel te verhogen, terwijl activatie van het kanaal spierontspanning lijkt te bevorderen. Hoewel dit specifieke aspect van het TREK-1 kanaal zich nog in een vroeg onderzoeksstadium bevindt, wordt het steeds belangrijker. Er bestaat hoop dat inzicht in de rol van moleculen zoals PE-22-28 bij spiercontractie en ontspanning nieuwe onderzoeksmogelijkheden kan bieden voor aandoeningen zoals myogene blaasdisfunctie en ook nieuwe wegen kan openen voor het begrijpen van de fysiologie van spierweefselontwikkeling.
Disclaimer
De genoemde producten zijn niet bedoeld voor menselijke of dierlijke consumptie. Research chemicaliën zijn uitsluitend bedoeld voor laboratoriumexperimenten en/of in-vitro testen. Elke vorm van lichamelijke toediening is strikt verboden volgens de wet. Alle aankopen zijn beperkt tot erkende onderzoekers en/of gekwalificeerde professionals. Alle informatie gedeeld in dit artikel is uitsluitend bedoeld voor educatieve doeleinden.
References
Djillani A, Pietri M, Moreno S, Heurteaux C, Mazella J, Borsotto M. Shortened Spadin Analogs Display Better TREK-1 Inhibition, In Vivo Stability and Antidepressant Activity. Front Pharmacol. 2017 Sep 12;8:643. doi: 10.3389/fphar.2017.00643. PMID: 28955242; PMCID: PMC5601071.
A. Djillani, J. Mazella, C. Heurteaux, and M. Borsotto, “Role of TREK-1 in Health and Disease, Focus on the Central Nervous System,” Front. Pharmacol., vol. 10, Apr. 2019, doi: 10.3389/fphar.2019.00379.
R. S. Duman, S. Nakagawa, and J. Malberg, “Regulation of adult neurogenesis by antidepressant treatment,” Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol., vol. 25, no. 6, pp. 836–844, Dec. 2001, doi: 10.1016/S0893-133X(01)00358-X.
H. Moha Ou Maati et al., “Spadin as a new antidepressant: absence of TREK-1-related side effects,” Neuropharmacology, vol. 62, no. 1, pp. 278–288, Jan. 2012, doi: 10.1016/j.neuropharm.2011.07.019.
Mazella J, Pétrault O, Lucas G, Deval E, Béraud-Dufour S, Gandin C, El-Yacoubi M, Widmann C, Guyon A, Chevet E, Taouji S, Conductier G, Corinus A, Coppola T, Gobbi G, Nahon JL, Heurteaux C, Borsotto M. Spadin, a sortilin-derived peptide, targeting rodent TREK-1 channels: a new concept in the antidepressant drug design. PLoS Biol. 2010 Apr 13;8(4):e1000355. doi: 10.1371/journal.pbio.1000355. PMID: 20405001; PMCID: PMC2854129.
C. Devader et al., “In vitro and in vivo regulation of synaptogenesis by the novel antidepressant spadin,” Br. J. Pharmacol., vol. 172, no. 10, pp. 2604–2617, May 2015, doi: 10.1111/bph.13083.
A. J. Silva, J. H. Kogan, P. W. Frankland, and S. Kida, “CREB and memory,” Annu. Rev. Neurosci., vol. 21, pp. 127–148,1998, doi: 10.1146/annurev.neuro.21.1.127.
Moha ou Maati, Hamid et al. “The peptidic antidepressant spadin interacts with prefrontal 5-HT(4) and mGluR(2) receptors in the control of serotonergic function.” Brain structure & function vol. 221,1 (2016): 21-37. doi:10.1007/s00429-014-0890-x
Q. Lei, X.-Q. Pan, S. Chang, S. B. Malkowicz, T. J. Guzzo, and A. P. Malykhina, “Response of the human detrusor to stretch is regulated by TREK-1, a two-pore-domain (K2P) mechano-gated potassium channel,” J. Physiol., vol. 592, no. 14, pp. 3013–3030, Jul. 2014, doi: 10.1113/jphysiol.2014.271718.
Alle bestellingen worden dezelfde dag verzonden indien geplaatst vóór 12:00 PST.
Dit product is uitsluitend bedoeld voor onderzoeks-/laboratoriumgebruik. Menselijk of dierlijk gebruik en/of consumptie is strikt verboden volgens de wet. Alleen gekwalificeerde en erkende professionals mogen deze producten hanteren. Alle informatie gevonden op Biotech Peptiden is uitsluitend bedoeld voor educatieve doeleinden. Raadpleeg onze algemene voorwaarden voor meer informatie.
Dr. Usman
Dr. Usman (BSc, MBBS, MaRCP) voltooide zijn studie geneeskunde aan het Royal College of Physicians in Londen. Hij is een enthousiaste onderzoeker met meer dan 30 publicaties in internationaal erkende peer-reviewed tijdschriften. Dr. Usman heeft gewerkt als onderzoeker en medisch consultant voor gerenommeerde farmaceutische bedrijven zoals Johnson & Johnson en Sanofi.



Reviews
There are no reviews yet.